(2)神経ネットワーク形成
上皮組織・臓器発生において、自律神経が重要な働きをしていることはよく知られている。肝臓の自律神経は、代謝、胆汁産生、血流量調整、組織修復や再生への関与が知られている。ヒトやモルモットでは実質領域にも神経線維が分布しているが、ラットではグリソン鞘内に限局して分布している。齧歯動物の実質領域では、肝細胞間にギャップ結合がよく発達しており、神経に代わって迅速な情報伝達を担っていると考えられている。しかしながら、肝臓の発生過程で、肝内神経網がどのように構築されるのかは分かっていなかった。
我々はマウスの発生過程における肝内の神経分布の変化を調べた。神経線維を同定するために、抗neural class III β-tubulin (Tubb3)を用いた。図1で示すように、成体マウス肝臓では、門脈周囲に微細な神経線維(白抜き矢頭)が見られ、グリソン鞘内のOsteopontin (OPN)陽性の胆管上皮細胞と肝動脈(HA)に隣接して発達している(黄矢頭)が、肝実質内には見られない。胎仔期(E17)から新生仔期(P1)までは、肝内に神経線維は見られず、肝外胆管・胆嚢に限局していた。生後1週になると、神経線維が肝門部のグリソン鞘内に認められるが、肝辺縁部では確認できない。
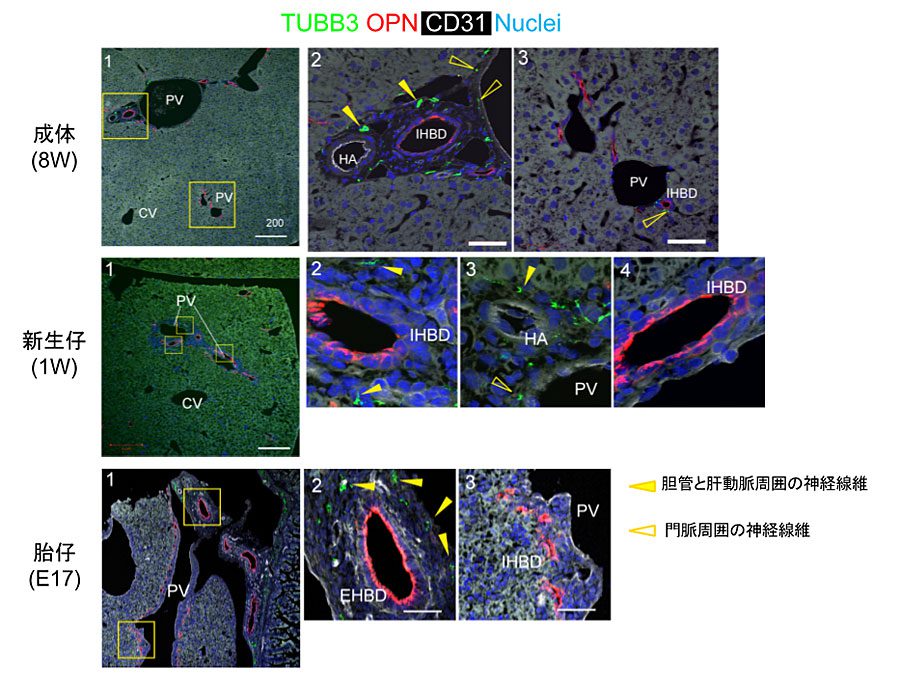 図1. マウス肝臓における神経の発達。PV: 門脈、IHBD: 肝内胆管、EHBD: 肝外胆管、HA: 肝動脈、CV: 中心静脈
図1. マウス肝臓における神経の発達。PV: 門脈、IHBD: 肝内胆管、EHBD: 肝外胆管、HA: 肝動脈、CV: 中心静脈
3次元的に神経線維の分布を解析すると図2で示すように、成体では大きな肝内胆管に沿って太い神経線維が走っており、辺縁部においても小葉間胆管に沿って神経線維が発達している。一方、生後1週目までは、成体と同様に大きな胆管や肝動脈に沿って神経は存在しているが細い線維からできている。この時期、まだ辺縁部まで神経線維は発達していない。
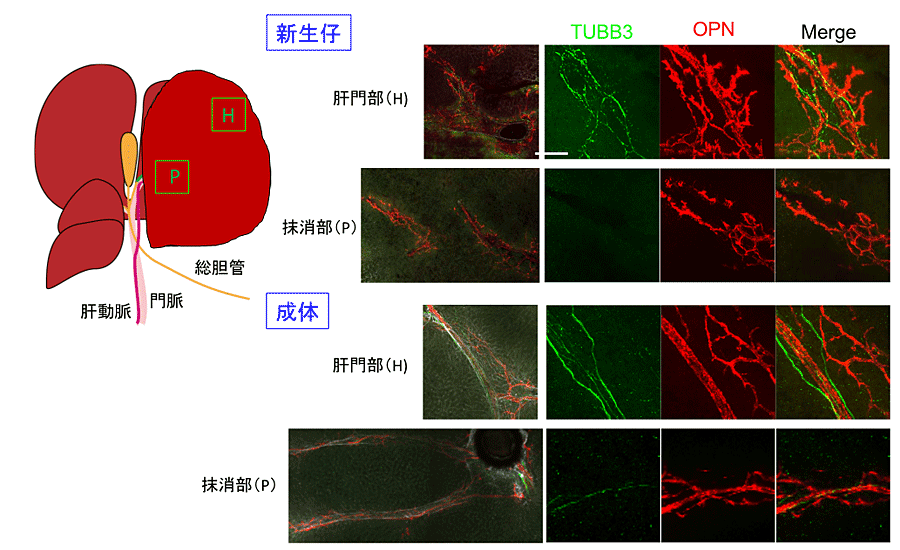 図2. 肝臓発生における神経ネットワークの形成。
図2. 肝臓発生における神経ネットワークの形成。
神経線維が辺縁部まで発達するのは生後3週目以降である(図3)。
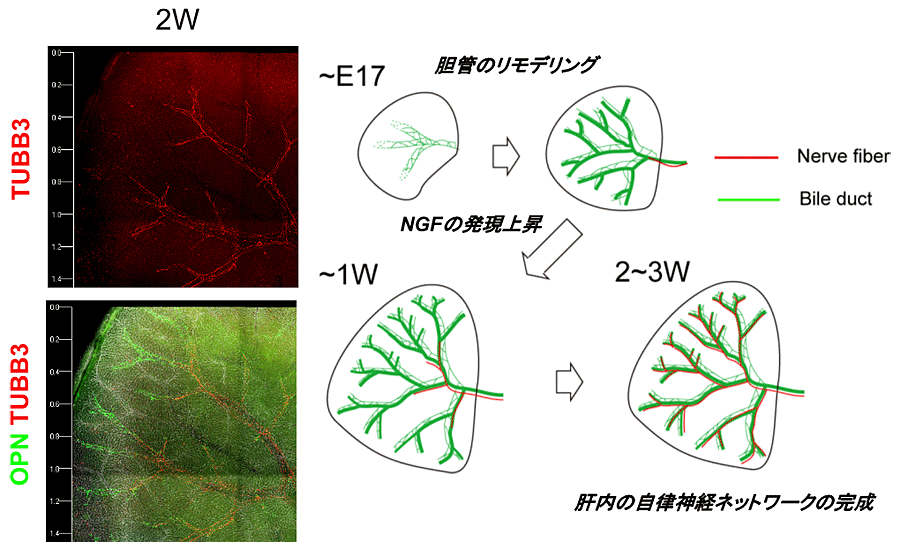 図3 肝内自律神経ネットワーク形成
図3 肝内自律神経ネットワーク形成
肝内胆管の発達に沿って神経ネットワークが形成されていることから、胆管上皮細胞が神経誘因因子を発現していることが推測された。新生仔期と成体の胆管上皮細胞の遺伝子発現を網羅的に調べた結果の解析から、胆管の発生に伴って神経成長因子であるNerve growth factor (Ngf)の発現が上昇することが分かった(図4)。さらにNGFの受容体であるTRKA (Ntrk1)が神経線維に認められた。NGFの発現は、胆管上皮細胞ばかりではなくThy1陽性間葉系細胞にも認められることから、肝内神経ネットワーク形成はグリソン鞘内の胆管上皮細胞と間葉系細胞が分泌するNGFによって誘導されることが予想された。
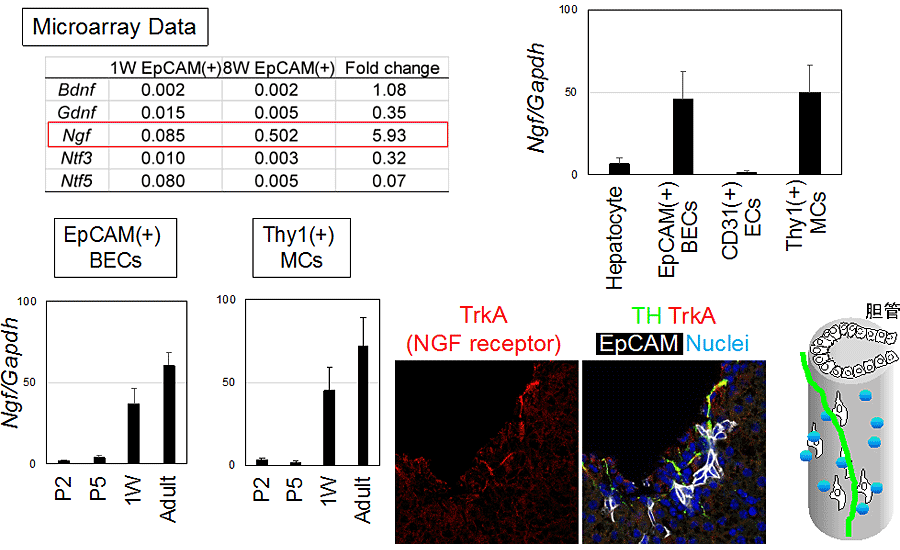 図4 神経成長因子Nerve growth factorとその受容体の発現
図4 神経成長因子Nerve growth factorとその受容体の発現
NGFが肝内の神経伸長を誘導する活性を持つことを検証するために、肝細胞にNGFを発現させてその影響を調べた。HTVi法によりNgf遺伝子を肝細胞に強制的に発現させると、小葉内に神経線維が伸長したことから、Ngfは肝内神経線維の伸長を誘導する因子であることが示された(図5)。
以上の事から、マウスにおいては、出生後に胆管でのNGF発現が上昇し、それに伴ってグリソン鞘に沿って神経線維が肝内組織に侵入することが明らかになった。
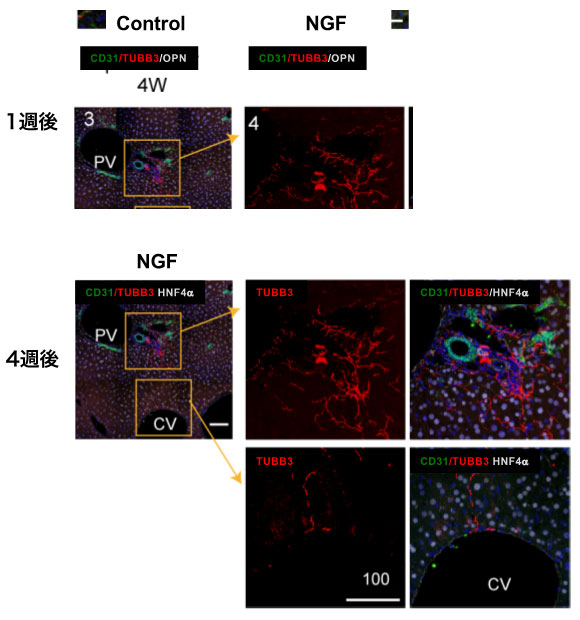 図5 NGF強制発現による肝小葉内神経誘導。
図5 NGF強制発現による肝小葉内神経誘導。
肝障害とその後の再生過程において、肝内神経ネットワークがどのような挙動を示すのかを調べるために、胆管細胞障害を起こすことが知られている4-diaminodiphenylmethane (DAPM)をマウスに投与して肝内胆管障害を誘導した。総胆管から逆行性に黒インクを注入した後、肝臓を透明化すると、肝内胆管の全体像を観察することができる。この手法を用いて胆管を可視化したところ、図6Aで示すように、DAPM投与後2日目ではインクが胆管から滲み出ており、小葉間胆管が障害されていることが分かる。組織学的には、小葉間胆管の消失と大きな肝内胆管の拡張が起こっていたが、小葉間胆管はDAPM投与後12日までには再生された。
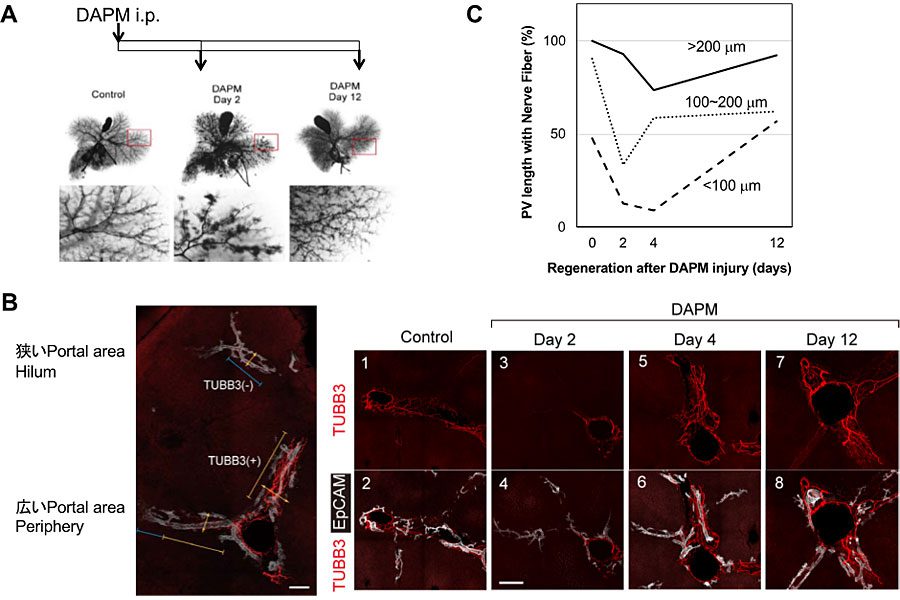 図6. DAPM投与による肝内胆管障害における神経再生。(A)胆管から逆行性に黒インクを注入した後、肝臓の透明化を行い肝内胆管を可視化した。(B)DAPM投与後の神経線維の再生。胆管上皮細胞(白;EpCAM+)、神経線維(赤;TUBB3)(C)門脈の長径と神経線維(TUBB3)の存在率の関係
図6. DAPM投与による肝内胆管障害における神経再生。(A)胆管から逆行性に黒インクを注入した後、肝臓の透明化を行い肝内胆管を可視化した。(B)DAPM投与後の神経線維の再生。胆管上皮細胞(白;EpCAM+)、神経線維(赤;TUBB3)(C)門脈の長径と神経線維(TUBB3)の存在率の関係
肝辺縁の小葉間胆管近傍の神経線維は、DAPM投与後2,4日目ではほぼ消失していた。門脈の大きさは肝臓内の位置と相関することから、門脈の大きさとその周囲の肝内胆管および神経線維の障害の程度を調べた(図6B, C)。肝門部に近い太い門脈があるところでは胆管構造が残存しており、神経線維の消失率も軽度であった。一方、、辺縁部の細い門脈(<100 μm)があるところでは胆管および神経線維が消失していた。つまり、胆管障害と神経線維の消失は、ともに肝門部より辺縁部で強く起こっていることが分かる。受傷後2日目に胆管上皮細胞は活発に増殖し、胆管上皮細胞と間葉系細胞、加えて血管内皮細胞にNgf遺伝子の発現上昇が見られた。
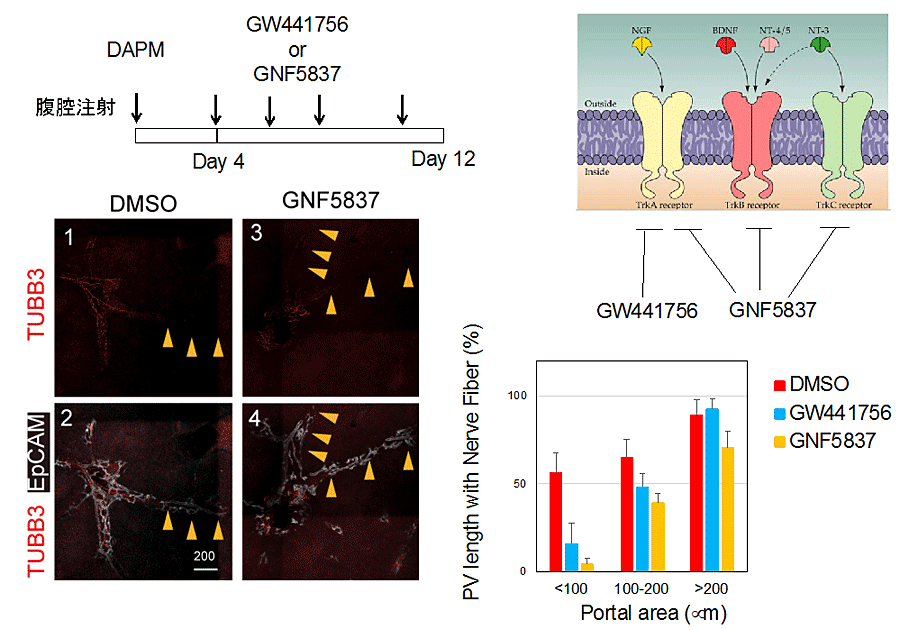 図7
図7
Ngfシグナルが神経再生に寄与しているか検討するために、受容体(Trk)阻害剤(GW4417566, GNF5837)を投与し、DAPM投与による胆管障害後の神経再生を調べると、胆管障害および神経消失が著しい辺縁部での再生が強く阻害されることが分かった。
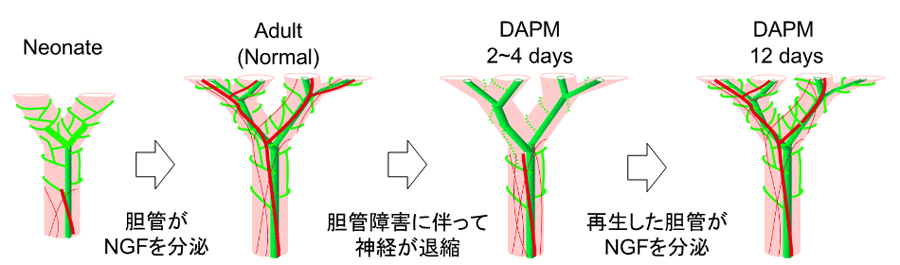 図8. DAPM投与による胆管障害後の神経再生
図8. DAPM投与による胆管障害後の神経再生
この結果をまとめると、図8で示すように、胆管障害起こるとNGFの分泌が減り、神経の退縮が起こる。その後、胆管再生に伴って分泌されるNGFにより神経再生が起こると考えられる。
結論
肝内の自律神経の発達は、肝内胆管のリモデリングがほぼ終わる1週目頃から、胆管上皮細胞と間葉系細胞の分泌するNGFにより誘導されて起こり、生後3週目頃には肝臓辺縁部まで達して完成する
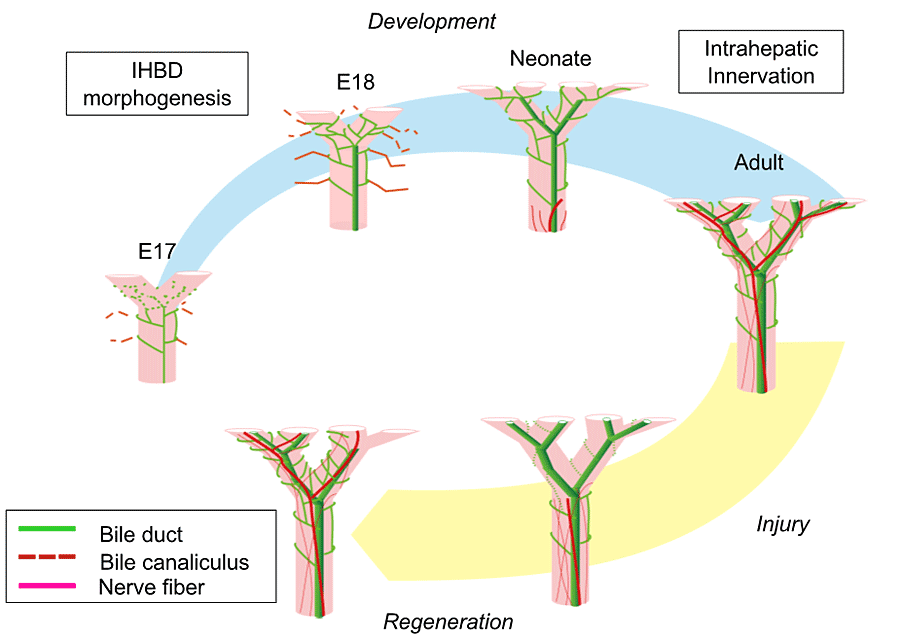 図9
図9
今後の課題
自律神経の活性が、部分肝切除後の肝再生に関与していることが知られており、Acetylcholineがマクロファージに作用してIL6発現を活性化することが報告されている。しかしながら、慢性的な肝疾患やその後の組織再生過程における自律神経の役割については不明であり、さらなる解析が必要である。
- Tanimizu N, et al. Intrahepatic bile ducts guide formation of the hepatic nervous system in developing mouse liver. Development, Apr 25; 145(9). pii: dev159095 (2018)
- Tanimizu N, et al. Intrahepatic bile ducts are developed through formation of homogeneous continuous luminal network and its dynamic rearrangement. Hepatology, 64(1):175-188 (2016)